La finalité objective en biologie
Prologue
Prologue
J’ai fait dans le passé plusieurs conférences sur la physique et les mathématiques. En physique, on se demande si l’on parviendra jamais au niveau le plus bas de l’infiniment petit de même qu’en astrophysique, on se demande si l’on atteindra jamais le niveau le plus élevé de l’infiniment grand. En mathématiques, on sait depuis Gödel que certaines questions sont indécidables. Personne n’a encore démontré la conjecture de Goldbach1 ni la convergence des grêlons du problème de Syracuse2, mais personne n’a encore découvert de contre-exemple.
Dans tous ces problèmes, les chercheurs se trouvent fréquemment confrontés à des systèmes complexes. Toutefois, cette complexité est du type que je qualifie de » simple » [13], car la structure des systèmes en question est connue et en permet donc l’analyse exhaustive. Au contraire, en biologie, la complexité est du type » compliqué « , même au niveau élémentaire de la cellule, en raison du nombre astronomique des éléments et de leurs interactions au sein de structures mal connues. Pour essayer de comprendre les phénomènes dont les systèmes vivants sont le siège, on est conduit à faire intervenir une certaine forme de finalité. C’est de cette finalité, que je qualifie » d’objective « , que je désire parler ce soir.
La cellule est l’élément constitutif de tous les êtres vivants. Sa complexité saute aux yeux, tant sont nombreux ses composants et leurs interactions. Chaque seconde, des milliers de réactions chimiques se déroulent au sein des dizaines d’organites présents dans chaque cellule, dont les produits sont véhiculés dans des sortes de sacs appelés vésicules par des milliers de molécules motrices sans cesse en mouvement, en un ballet qui semble au premier abord désordonné, mais est le signe visible d’une activité hautement coordonnée d’échanges moléculaires ayant chacun un rôle spécifique dans le métabolisme cellulaire. Le plus extraordinaire est que les biologistes ont réussi, avec une patience et une persévérance en tous points remarquables, à mettre en évidence, dans ce maquis qui semble impénétrable, quelques processus fondamentaux qui permettent d’expliquer les mécanismes de l’hérédité et de l’embryogenèse.
La finalité objective
C’est un fait que l’homme est aujourd’hui sur la Terre et qu’il est le seul être vivant à saisir l’Univers dans son ensemble, dans l’espace comme dans le temps. Quand on récapitule les étapes de l’évolution cosmique et biologique (figure 1), force est de constater que l’homme en est l’aboutissement actuel, peut-être provisoire. Il existe un fossé infranchissable entre les animaux les plus évolués et l’homme moderne. Ce hiatus est dû à la main et, surtout, au langage, qui permet l’accumulation des connaissances et leur transmission de génération en génération. Des événements contingents, tels que des chutes de météorites ou des éruptions volcaniques, sont intervenus dans le déroulement de l’évolution ayant conduit à l’apparition de l’homme, qui aurait pu ne jamais apparaître. Mais c’est un fait qu’il est apparu et qu’il est devenu le maître de la Terre, pour le meilleur et pour le pire, conformément aux Écritures.
Ces considérations, qui ne font que constater un état de fait, conduisent à l’idée d’une » finalité objective « , l’adjectif » objectif » se rapportant à un fait que l’on ne peut que constater sans pouvoir l’expliquer. L’histoire de la Seconde Guerre mondiale m’en fournit un exemple saisissant. Après Stalingrad, l’armée allemande s’est comportée comme l’alliée objective des forces occidentales, en s’opposant à la poussée vers l’Ouest du rouleau compresseur soviétique qui, en cas d’effondrement de l’armée allemande, aurait atteint Brest bien avant le débarquement allié en Normandie. Je pense que cet exemple historique fait bien comprendre le sens que je donne à l’adjectif » objectif « .
On sait que Newton n’aimait pas l’idée de la transmission instantanée d’une force à distance. Il exprimait sa loi de gravitation universelle en disant : » Tout se passe comme si les corps s’attiraient en raison directe de leur masse et en raison inverse du carré de leur distance. » Faut-il l’imiter et dire : » Tout se passe comme si l’homme était la fin de l’Univers » ? Cette formulation ne peut pas être accusée de finalisme ; c’est la simple constatation d’une finalité objective. Elle ne peut être identifiée à la forme forte du principe anthropique, justement parce que ce n’est pas un principe.
Déterminisme et finalité
Les actions humaines ont toujours un but, c’est-à-dire un objectif fixé dans un avenir plus ou moins lointain. Avant d’agir, l’homme réfléchit. Il se dit que ce qui s’est passé hier a toutes chances, toutes choses égales d’ailleurs, de se reproduire demain. Autrement dit, il se place dans une optique déterministe. Si le monde n’était pas en gros déterministe, toute action raisonnable serait impossible.
Dans un monde parfaitement déterministe, la causalité et la finalité se rejoignent, puisqu’on peut indifféremment suivre l’évolution d’un phénomène de son état initial à son état final ou, inversement, remonter de l’état final à l’état initial. La prédiction et la rétrodiction sont également possibles, la rétrodiction étant la prédiction de l’état initial à partir de l’état final.
La cybernétique
| Figure 1 | |
| Événements |
Dates (millions d’années dans le passé) |
| Big bang Galaxie Système solaire Terre Premières molécules autoreproductrices Bactéries, algues bleues Photosynthèse Eucaryotes Oxygène Méduses, coraux Coquillages, crustacés Vertébrés, explosion cambrienne Plancton, trilobites Poissons, plantes terrestres Insectes Amphibiens Reptiles Dinosaures Petits mammifères, extinction permienne (90%) Oiseaux Fleurs Cétacés, primates Extinction (80 %), fin des dinosaures Grands mammifères Pongidés, hominidés Homo habilis Homo sapiens Peintures rupestres |
15 000 14 000 – 10 000 5 000 4 500 4 000 3 500 2 000 1 800 1 300 1 000 800 600 550 500 450 400 350 300 250 200 150 100 65 40 5 – 10 1,5 – 2,5 0,3 0,03 |
|
Principales dates de l’évolution cosmique et biologique. On note l’accélération du processus. Le rapport entre l’âge de l’Univers visible et la date des événements est de 3 pour la Terre, de 25 pour les vertébrés, de 150 pour les primates et de 50 000 pour l’homo sapiens. |
|
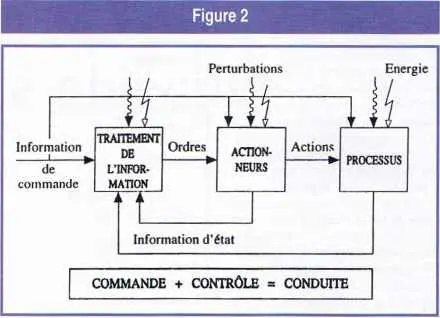
La finalité est donc inscrite dans tous les processus d’action intentionnelle, dont la théorie et la pratique s’appellent la cybernétique. Ce mot, repris par Norbert Wiener de la classification des sciences d’Ampère, où il désignait l’art de gouverner, a été défini par lui comme la » théorie de la commande et de la communication chez l’animal et dans la machine « . C’est assez dire sa généralité. On peut parler aussi de théorie et pratique des processus d’action orientés vers un but. On peut voir sur la figure 2 le schéma général de tous les processus d’action.
La cybernétique est donc la science des processus gouvernés par une finalité. Depuis cinquante ans, elle a profondément imprégné les réflexions, non seulement des ingénieurs, mais aussi des biologistes, qui ont compris l’importance des rétroactions dans les réseaux cybernétiques de la matière vivante. On se demande pourquoi, dans ces conditions, la finalité a aujourd’hui si mauvaise presse parmi les biologistes. Ils l’acceptent pour expliquer le fonctionnement régulé des organes tels que le foie ou le cœur, mais la rejettent dès qu’elle semble remettre en cause les sacro-saints principes du darwinisme.
Mais, en pratique, le langage des biologistes est toujours finaliste. Ils nous expliquent que les yeux sont faits pour voir, les ailes pour voler et l’estomac pour digérer. Jacques Monod a tellement peur d’être taxé de finaliste qu’il invente le mot » téléonomie » pour désigner la propriété de la matière vivante de réaliser un projet, c’est-à-dire de répondre à une finalité. Or, pour réaliser un projet, il faut que ce projet soit décrit par un plan et, de plus, qu’il y ait un projeteur. Le mot » téléonomie » est donc beaucoup plus fort que l’expression » finalité objective « , que je lui préfère.
Les bifurcations
Les bifurcations introduisent dans le déroulement déterministe d’un processus des éléments contingents qui interdisent la prédiction. C’est le cas d’un torrent qui dévale le flanc d’une montagne et qui rencontre une protubérance qui le fait dévier vers la droite ou vers la gauche sous l’effet d’une petite perturbation imprévisible. L’existence de cette bifurcation empêche de prévoir l’état final, mais elle n’interdit pas la rétrodiction, puisque les deux branches issues du point de bifurcation se rejoignent en amont. La rétrodiction demeure déterministe alors que la prédiction ne l’est plus.
Cela n’altère pas le finalisme global du phénomène, puisque l’eau recherche toujours son état d’énergie potentielle minimale en rejoignant, par un chemin qui n’est pas entièrement déterminé, le fleuve ou le lac situé en contrebas, sans qu’on puisse lui en prêter l’intention.
Les principes d’extremum
Schéma général des processus d’action. La comparaison de l’information de commande et de l’information d’état dans l’organe de traitement de l’information permet d’élaborer les ordres qu’il convient de donner aux actionneurs agissant sur le processus de manière à ramener celui-ci vers son régime assigné en dépit des perturbations extérieures.
Le raisonnement qui vient d’être fait s’applique-t-il à tous les principes d’extremum, tels que le principe de Fermat ou le principe de moindre action ? Cela dépend du point de vue où l’on se place.
On peut les considérer comme une propriété du modèle mathématique du phénomène étudié. Mais on peut aussi leur donner une interprétation statistique en appliquant la méthode de l’intégrale des chemins de Richard Feynman. Considérons le trajet d’un photon d’un point A à un point B et faisons la somme de toutes les trajectoires possibles joignant A à B. Cette somme est une somme vectorielle dont chaque élément possède une amplitude et une phase. Loin de la droite reliant A et B, les phases présentent toutes les valeurs possibles, de sorte que les contributions des diverses trajectoires s’annulent. Elles s’ajoutent au contraire au voisinage de cette droite, où les phases sont voisines, et l’on retrouve la propagation rectiligne de la lumière.
Ce raisonnement s’applique à tous les phénomènes de l’optique géométrique, qui se trouvent ainsi reliés à la physique quantique. Il réconcilie le principe finaliste du moindre trajet avec le déterminisme.
Qu’est-ce que la vie ?
On ne sait pas définir la vie ; on peut seulement énumérer les propriétés qui caractérisent les êtres vivants. En bref, on peut dire que les êtres vivants naissent, se nourrissent, croissent, se protègent, se reproduisent et meurent. Il n’y a pas de cycle plus déterministe que celui de la vie, car il ne souffre aucune exception. Ce faisant, les êtres vivants interagissent sans cesse avec leur environnement et avec leurs semblables.
La vie est une propriété d’une structure plutôt que d’un objet particulier. Les cellules d’un être vivant sont remplacées à plusieurs reprises. C’est leur organisation qui est importante. C’est l’apologue de la barque de Delphes, dont toutes les planches ont été remplacées les unes après les autres, mais qui reste toujours la même barque. La vie implique un mécanisme d’autoreproduction, qui suppose que l’être vivant contient une description de lui-même.
Cette description est contenue dans les chaînes d’acides nucléiques constituant l’ADN présent dans les noyaux de toutes les cellules. Un être vivant possède un métabolisme, qui convertit la matière et l’énergie de l’environnement dans les formes utiles à l’organisme. Un être vivant est séparé de son environnement par une membrane permettant des échanges sélectifs dans les deux sens. Ses composants sont de même isolés par des membranes.
L’unité du vivant est devenue évidente avec les progrès de la biologie moléculaire, qui montre l’universalité de l’ADN, présent chez tous les êtres vivants, et du code génétique qui définit la correspondance entre les codons de l’ADN et les protéines. Cette universalité renforce l’idée que tous les êtres vivants, de la bactérie à l’homme, ont une origine commune dont ils dérivent par le mécanisme de l’évolution.
La finalité de la vie
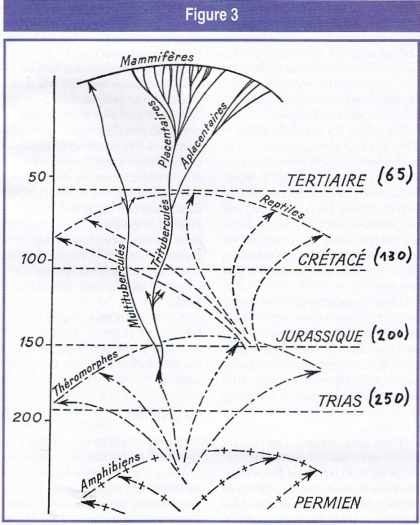
Schéma imaginé par Pierre Teilhard de Chardin pour symboliser le développement en nappes des tétrapodes. Les chiffres à gauche expriment des millions d’années. J’ai mis à droite, entre parenthèses, des chiffres plus conformes aux connaissances actuelles, mais on voit qu’ils diffèrent peu de ceux de Teilhard (figure empruntée au Phénomène humain).
On peut considérer que la finalité de la vie réside dans son maintien par la reproduction dans les pires conditions. J’ai déjà cité l’exemple de la grande douve [13] qui n’atteint sa destination dans le foie du mouton, sous sa forme adulte, qu’en passant par cinq stades larvaires, avec une énorme perte de larves. J’ai aussi cité le tardigrade, animal transparent d’un millimètre de long, à pattes non articulées, qui est capable de se déshydrater complètement et d’arrêter son métabolisme afin de supporter des conditions extrêmes de température, de pression et de radiations. Il peut ainsi rester en léthargie, par exemple dans les glaces polaires, et reprend vie dès qu’il se retrouve en présence d’eau liquide.
Dans les fosses marines des Galapagos vit une faune extrêmement riche qui comporte des vers de 2 m de long colonisés par des bactéries, des crabes, des crevettes, des poissons, des moules géantes, du plancton et des prédateurs tels que des poulpes. Cette vie se maintient dans des sources chaudes d’origine volcanique, à des températures pouvant atteindre 350 °C. En l’absence de lumière, la photosynthèse y est remplacée par la chimiosynthèse, qui fabrique la matière organique à partir du gaz carbonique dissous en puisant son énergie dans les composés soufrés contenus dans l’eau chauffée par le magma.
De même pendant les glaciations, toute forme de vie disparaissait de la surface gelée de la Terre. Mais, dès que les conditions climatiques devenaient plus clémentes, la vie reprenait ses droits dans un surprenant renouveau. Cette forme de finalité, qui fait que la vie se maintient dans les pires conditions, est un fait d’observation que nul ne peut contester. C’est la finalité objective par excellence. C’est » l’élan vital » d’Henri Bergson. La vie renaissante occupe toutes les niches, même les moins prometteuses.
Ainsi, on peut retourner le propos darwinien et dire que seuls les êtres vivants qui ne sont pas du tout adaptés à leur milieu ne subsistent pas !
L’évolution biologique
Le néodarwinisme
Le néodarwinisme pose en principe que l’évolution biologique résulte de l’action combinée des mutations aléatoires et de la sélection naturelle. Les mutations se produisent au hasard et entraînent des modifications de certains caractères qui peuvent être avantageuses ou non pour la survie de l’espèce, c’est-à-dire pour la fécondité des générations successives. Les mutations désavantageuses disparaissent donc avec les individus qui les portent et se reproduisent mal, tandis que les mutations avantageuses se retrouvent chez un nombre croissant de descendants. Tout se passe comme si les caractères correspondants étaient héréditaires. En outre, l’apparition de caractères vraiment nouveaux ne constituant pas de simples variantes se fait progressivement par l’effet cumulatif de micromutations qui se renforcent mutuellement et finissent par produire des effets macroévolutifs.
La stabilité des espèces
À l’intérieur d’une même espèce, la finalité ne fait aucun doute : un gland donne toujours un chêne et un œuf de tortue une tortue. Cette finalité se traduit par l’extraordinaire stabilité des espèces, dont certaines subsistent, inchangées, depuis 500 millions d’années. Le plan de l’organisme adulte est donc manifestement contenu dans l’œuf ou dans la graine. Dans ces conditions, l’adaptation à un environnement changeant est bien expliquée par l’accumulation des effets des micromutations sous la pression sélective.
Des exemples classiques sont les pinsons observés par Charles Darwin aux Galapagos, dont la forme du bec évolue en fonction de la nourriture disponible dans les différentes îles, et les phalènes du bouleau dont la population s’assombrit dans une atmosphère charbonneuse, dans laquelle les phalènes clairs sont plus aisément repérables par les oiseaux. Cette dernière transformation est réversible et les phalènes s’éclaircissent lorsque la pollution en poussière de charbon diminue. Mais ces adaptations n’ont pas créé d’animaux autres que des pinsons et des phalènes.
La spéciation
Il est plus difficile d’expliquer par l’effet cumulatif de petites mutations l’apparition des amphibiens à partir des poissons ou des oiseaux à partir des reptiles. Les archives fossiles manquent singulièrement de formes intermédiaires montrant clairement la transition entre deux espèces censées dériver l’une de l’autre. Les exemples tels que celui des premiers chevaux, qui avaient la taille d’un petit chien, sont rares.
Il y a soixante-dix ans, dans Le phénomène humain, Pierre Teilhard de Chardin avait déjà donné de l’absence de fossiles d’espèces intermédiaires une explication assez convaincante. J’ai reproduit sur la figure 3 le schéma qu’il avait imaginé pour symboliser le jaillissement des espèces à partir d’un ancêtre issu d’une classe déjà stabilisée.
Cette vision teilhardienne a été reprise par Gould et Eldridge dans leur théorie des isolats périphériques. Considérons, par exemple, l’émergence des reptiles à partir des théromorphes, animaux intermédiaires entre les amphibiens et les reptiles, dont font partie les thérapsides et les tortues. Des mutations se produisent dans un petit groupe isolé de théromorphes. Ces mutations produisent un grand nombre de variantes, dont seules un petit nombre » réussissent « , tandis que la plupart s’éteignent. Toutes ces variantes sont des animaux de petite taille, encore assez mal définis et mal ossifiés, et peu nombreux. Ils n’ont pas laissé de fossiles. Les lignées divergentes s’isolent en une gerbe close, imperméable aux gerbes voisines. Leurs éléments terminaux, stabilisés, se prolongent jusqu’à nous par leurs survivants et leurs fossiles. Les transitions se produisant en des temps » courts » à l’aune de l’évolution, tandis que les périodes stables, appelées aujourd’hui stases, peuvent durer des millions d’années.
De même, les premiers mammifères, apparus alors que dominaient les dinosaures, étaient petits et souvent fouisseurs. Ils ont pu résulter de mutations survenues dans des populations de petits reptiles. On peut imaginer que des femelles n’ont pas pondu leurs œufs et que ceux-ci ont éclos en elles. Ainsi seraient apparus des ébauches de marsupiaux, puis d’animaux placentaires.
La sélection naturelle joue dans le tri des variantes créées par tâtonnements, non dans la spéciation proprement dite. Par suite de ce mécanisme il ne subsiste aujourd’hui qu’un petit nombre des espèces ayant existé dans un passé lointain et beaucoup de formes éteintes nous demeureront inconnues.
Dans cette optique, l’archéoptéryx ne serait pas une forme intermédiaire entre les reptiles et les oiseaux, mais une forme accomplie de reptile volant, qui a dû disparaître avec les dinosaures terrestres il y a soixante-cinq millions d’années. Ce serait des animaux arboricoles plus petits, qui n’ont pas laissé de traces, qui auraient, après de multiples tâtonnements, donné naissance aux oiseaux que nous connaissons.
La complexification
Le darwinisme n’explique pas la tendance générale des nouvelles espèces à devenir de plus en plus complexe. Cette complexification entraîne la croissance et la centralisation du système nerveux, car la coordination des éléments d’un organisme complexe entre eux et avec l’environnement nécessite un nombre croissant de neurones et la création d’aires de traitement centralisé des signaux neuronaux dans un organe spécialisé, le cerveau. La complexification et la céphalisation qu’elle entraîne peuvent constituer des avantages dans la lutte pour la survie de l’espèce, en améliorant le traitement des signaux reçus du monde extérieur. Mais les bactéries, qui existent depuis 3,5 milliards d’années, sont parfaitement bien adaptées et n’ont jamais manifesté la moindre tendance à se complexifier.
D’autre part plus une machine est complexe, plus elle présente de risques de mauvais fonctionnement. De même, chez les êtres vivants, la multiplication et la complexification des organes augmentent les risques de maladies dues notamment aux agressions microbiennes. C’est pourquoi le système immunitaire se développe avec la complexité. Pourquoi la Nature a‑t-elle choisi la voie de la complexification croissante, qui est aussi celle de la fragilisation ?
On ne peut donc, ici encore, qu’imiter Newton et dire : » Tout se passe comme si l’évolution biologique était orientée dans le sens de la complexification et de la céphalisation. »
Le génome
Il a été dit plus haut que l’œuf ou la graine contenaient le plan de l’organisme. On peut penser que ce plan est matérialisé par le génome présent dans chacune des cellules de cet organisme et dont les gènes s’expriment d’une manière hautement coordonnée au cours de son développement.
Le génome est l’ensemble des gènes d’un organisme, un gène étant un fragment d’ADN définissant une protéine par la suite de ses acides aminés. La synthèse des protéines se fait en deux temps, la transcription d’un fragment d’ADN en ARN messager dans le noyau cellulaire, suivie de la traduction, au cours de laquelle chaque triplet d’ARN messager donne dans le ribosome un triplet correspondant d’ARN de transfert associé à un acide aminé qui est censé être disponible dans le cytoplasme. La protéine résulte de la concaténation de ces acides aminés ; ses propriétés résultent de son mode de repliement.
Cependant, le texte génomique ne suffit pas pour définir la construction et le fonctionnement de la cellule et de l’organisme. En effet, l’expression des gènes contenus dans une cellule dépend de sa localisation dans l’organisme, du stade de son développement et des messages qu’elle reçoit des cellules voisines et de l’environnement. On ne peut donc pas dire que le génome définit le plan de l’organisme, puisque la réalisation de ce plan dépend des conditions qu’elle crée dans l’organisme en construction. L’expression des gènes est gouvernée dans l’espace et dans le temps par l’organisme qu’ils édifient.
De plus, les parties codantes de l’ADN ou exons sont séparées par des parties non codantes ou introns qui doivent être excisées pendant la transcription, de sorte qu’un gène peut être transcrit par segments qui sont coupés et regroupés par la chimie cellulaire. Mais ce regroupement peut se faire de différentes manières et donner naissance à une variété d’ARN messagers différents définissant donc des protéines différentes.
Enfin, les protéines ainsi synthétisées peuvent être modifiées par des facteurs externes tels que température et lumière et se replier différemment sous l’influence de molécules » chaperons « . Une même protéine peut ainsi avoir plusieurs rôles différents.
D’autre part, quand on connaît le génome complet, on ne connaît pas pour autant les gènes, surtout s’il y a beaucoup d’introns qui ne sont pas forcément formés de triplets. La localisation des limites d’un gène est toujours une opération difficile, car on trouve un peu partout des signaux de début et de fin de lecture. On est donc obligé d’essayer toutes les manières de lire la séquence et de la décomposer en triplets, la traduire en protéines grâce au code génétique et comparer le résultat avec le contenu de banques de données pour rechercher des protéines similaires. » C’est très compliqué et il y a beaucoup d’erreurs » (A. Danchin). Il faudra beaucoup de temps pour identifier tous les gènes contenus dans un génome et plus encore pour déterminer leurs rôles.
Quant au génome humain, il contient entre 30 000 et 100 000 gènes. Sa partie codante représente 3 % du génome et contient 100 millions de paires de bases. Le reste est déclaré inutile, mais qu’en sait-on ? Si l’on séquence une partie du génome on a peu de chances d’y trouver la partie codante d’un gène. De plus, la structure fine du génome humain est très polymorphe, un jeu de chromosomes provenant du père et l’autre de la mère et les gènes homologues pouvant être des allèles différents. La séquence qui résulte des programmes de séquençage est très ambiguë. Si l’on séquence les gènes de plusieurs individus, on obtient un patchwork rassemblant des séquences parfaitement incompatibles, car les différents allèles qu’elles contiennent ne peuvent coexister.
En résumé, on peut admettre que le génome contient le plan de l’organisme, à condition de préciser que c’est un plan dynamique, dont la réalisation dépend de son déroulement. Un même gène ne s’exprime pas de la même manière dans les différentes étapes de l’embryogenèse et ne fabrique donc pas nécessairement la même protéine.
Épilogue
On pourrait considérer la finalité comme une propriété immanente de l’Univers dans la mesure où l’homme considère celui-ci comme un tout cohérent soumis à des lois universelles. Chacun de ses composants aurait alors sa propre finalité, qui serait de contribuer à la cohérence de l’ensemble en se comportant de manière que les lois soient vérifiées. L’homme est l’un de ces composants et on ne peut donc pas donner une image cohérente de l’Univers sans faire référence explicite à son existence. On rejoint ainsi le principe anthropique sans sortir de la science, en s’abritant derrière un modèle idéal de 1’Univers. Malheureusement, un tel modèle n’existe pas encore et n’existera peut-être même jamais. La finalité immanente de l’Univers est donc tout compte fait une finalité objective et rien de plus.
On peut résumer ce qui a été dit à propos de la finalité objective de l’évolution biologique en disant que tout se passe comme si celle-ci était orientée dans le sens de la complexité, qui va de pair avec la céphalisation et, partant, avec la montée de la pensée et de la conscience. On retrouve ainsi ce que Teilhard de Chardin appelait la » loi de complexité-conscience « .
Le darwinisme est décidément une théorie trop simpliste pour rendre compte de la complexité du monde vivant passé et présent. Non seulement il n’explique pas les orientations manifestes de l’évolution biologique, mais il n’explique pas non plus l’extraordinaire stabilité de certaines espèces d’insectes et de crustacés dont certaines, telles que les limules, subsistent, inchangées, depuis cinq cents millions d’années.
Tous ces phénomènes traduisent la tendance irrépressible au maintien de la vie par les stratégies les plus invraisemblables et à n’importe quel prix. C’est l’élan vital dont parlait Henri Bergson, qui est en fin de compte l’expression pratique de la finalité objective de la vie, qui vise avant tout à son maintien.
______________________________
1. La conjecture de Goldbach s’énonce en disant que tout nombre pair peut s’écrire sous la forme de la somme de deux nombres premiers. Exemple 52 = 23 + 29.
2. La suite des nombres de Syracuse s’obtient, à partir d’un impair quelconque N, en calculant (3 N + l), en le divisant par 2 autant de fois que possible et en réitérant cette opération. Les nombres obtenus oscillent en montant et en descendant comme les grêlons dans un nuage d’orage, mais on n’a jamais trouvé de suite qui ne finisse pas par descendre vers 1, ou plutôt vers le cycle 1, 4, 2, 1…

