Structures dans l’Ensemble de Mandelbrot
J’ai découvert l’Ensemble dans les articles de Dewdney, mais faute d’ordinateur personnel je n’en ai rien tiré à l’époque. C’est la rencontre fortuite avec le programme Chaos qui a relancé mon intérêt en 1993, et cette fois j’ai décidé de ne pas en rester à l’examen aléatoire d’images belles mais incompréhensibles.
Adoptant l’approche méthodologique de Mandelbrot, et m’appuyant sur des mathématiques de taupe accessibles à tous, j’ai poussé l’analyse des images bien au-delà de tout ce qui avait été fait jusqu’à présent, et trouvé les lois, extrêmement simples et générales, qui régissent la structure de l’Ensemble.
Cette découverte, vérifiée sur un très grand nombre de cas (littéralement plus de 10 000), reste une simple conjecture car je n’ai pas trouvé de démonstration. J’espère convaincre les mathématiciens de s’y intéresser.
En attendant, elle permet à tout un chacun, utilisant le programme Chaos et quelques additions de mon cru, de comprendre la structure de n’importe quelle image, et d’en prédire quantitativement les caractéristiques, jusqu’aux plus infimes détails accessibles (grossissement 1014 environ).
Rappels
Pour situer les découvertes, il faut rappeler ce qui était connu en 1993 en matière de structure de l’Ensemble. J’essaye d’être bref, mais il faut que la suite soit compréhensible.
Définition
L’Ensemble de Mandelbrot est l’ensemble des points C du plan complexe qui font converger l’itération :
Zn+1 = Zn2 + C, avec Zo = 0.
On démontre aisément que l’Ensemble est symétrique par rapport à l’axe horizontal, et intérieur au cercle de rayon 2 centré sur l’origine. On définit alors la divergence comme le franchissement de ce cercle.
L’algorithme de Mandelbrot consiste à colorier en noir les points C de l’Ensemble, et en couleurs les points qui conduisent à une divergence. Une couleur spécifique est associée à chaque valeur possible (plafonnée arbitrairement pour limiter la durée des calculs) du nombre d’itérations effectuées au moment de la divergence. Le choix de cette correspondance présente un grand intérêt sous l’angle esthétique, évidemment, mais aussi pour les mathématiciens. Le choix que j’ai fait dans Chaos établit un net contraste à la frontière de l’Ensemble, marquée par des couleurs claires : il est primordial pour la compréhension des structures.
Domaines
 La présentation des résultats à l’échelle 1 (le cercle de rayon 2 remplissant l’écran) met en évidence trois domaines dont l’étude mathématique révèle les spécificités (figure 1).
La présentation des résultats à l’échelle 1 (le cercle de rayon 2 remplissant l’écran) met en évidence trois domaines dont l’étude mathématique révèle les spécificités (figure 1).
Cette zone délimitée par une cardioïde, dont le point de rebroussement est à + 0,5 et le sommet à – 0,75, est le domaine de convergence vraie : à chaque valeur de C correspond une limite A que l’on sait calculer directement. Le contour de la cardioïde appartient au domaine. L’origine est sa propre limite.
À l’extérieur de la cardioïde la convergence change de nature : il n’y a pas une limite, mais un groupe de p valeurs formant cycle, dont p est la période. À chaque valeur de C correspond un jeu unique de valeurs et une période spécifique.
Le domaine 2 se compose d’une infinité de sous-domaines d’apparence circulaire, tangents à la cardioïde ou entre eux. À l’intérieur d’un sous-domaine, la période est la même en tous points, et j’appelle poids du sous-domaine cette période commune.
Je baptise » cercles » ces sous-domaines, bien que la preuve de circularité n’existe que pour le gros cercle axial ; des mesures sur certains autres montrent que l’écart à la circularité, s’il existe, est inférieur au dix millième. On peut classer ces cercles comme suit :
a) une famille primaire, symétrique par rapport à l’axe horizontal, commençant avec le cercle axial de poids 2 (noté C2), et se poursuivant avec des paires de cercles de poids 3 (C3, en haut et en bas), 4 (C4, à 45° du côté droit), 5, 6 (C6, tangents à droite de la cardioïde), etc., jusqu’à l’infini dans le point de rebroussement.
b) entre deux cercles primaires quelconques de poids p et p + 1 existe une série secondaire, avec un cercle secondaire majeur situé à peu près au milieu, de poids 2 p + 1, et deux séries infinies M et P qui l’encadrent. Par exemple, il existe une paire de CS5 à 45° entre C2 et C3, pour p = 2.
c) on définit ensuite de la même manière, et on observe, des séries tertiaires, quaternaires, quinaires, etc., jusqu’à l’infini. Le poids de chacun de ces cercles s’évalue de proche en proche en fonction du p initial, par simple addition des poids des cercles encadrants. Tous les cercles compris entre Cp et Cp + 1 ont ainsi un poids m de la forme a p + b : nous découvrirons plus loin l’importance du coefficient a.
On sait calculer les coordonnées du point de contact de chacun de ces cercles avec la cardioïde, et la tangente en ce point, qui sépare le domaine de poids 1 du sous-domaine de poids m.
Autour de chacun de ces cercles de premier niveau existe une hiérarchie de familles subordonnées, dont le numéro se calcule comme les poids ci-dessus. Le poids de chacun de ces cercles de 2e niveau est le produit de son numéro par le poids du cercle porteur.
Cette propriété est récursive, et il existe des cercles de 3e, 4e niveau, etc., jusqu’à l’infini. Sur l’axe, par exemple, il existe une infinité de cercles de poids 2, 4, 8, 16, etc.
Des formules existent pour déterminer le point de contact de chaque cercle avec son porteur. Cependant, seul le cas du porteur C2 est accessible en bloc à l’ordinateur. Pour les autres, le calcul est numérique et point par point.
À l’extérieur des domaines 1 et 2 jointifs, on observe des points noirs de convergence, en apparence non connectés. En grossissant ces points, on constate que chacun d’eux est une copie approximative de l’ensemble complet : on démontre que la correspondance est un homéomorphisme, qui conserve toutes les propriétés décrites ci-dessus, tout en autorisant des déformations, notamment en torsion.
On a démontré que l’Ensemble est connexe. Il faut donc trouver la structure qui intègre les microcardioïdes du domaine 3 avec les domaines 1 et 2.
Le nombre des objets de poids p, cercles ou microcardioïdes, est borné par le nombre 2p‑1 – 1. Mais, puisque p peut croître indéfiniment, le nombre des cercles et le nombre des microcardioïdes de l’ensemble sont tous deux infinis.
Rosaces
Une image de l’Ensemble (figure 1) montre la zone noire entourée d’une frange colorée. En grossissant un point quelconque de cette frange, on découvre qu’elle est constituée de dessins disjoints, chacun émanant d’un cercle.
Une propriété remarquable, non démontrée mais générale, est que le dessin issu d’un cercle de poids p est une rosace à p bras, dont un pédoncule rattachant le cœur de la rosace au cercle. Nous notons B1 le pédoncule, B2 à Bp les bras libres.
Chaque bras libre sert lui-même de B1 à une rosace terminale Rp à p bras, et cela récursivement.
On constate que les microcardioïdes sont exclusivement localisées dans les bras, ce qui suggère que la topologie de l’Ensemble est en étoile.
Les textes mathématiques ne parlent que peu de ces dessins, qu’ils baptisent « antennes », de sorte qu’il est difficile de séparer ce qui était connu avant que j’étudie ces dessins, de ce qui ne l’était pas. Je dirai seulement que je n’ai trouvé dans aucun texte une quelconque allusion à la structure interne des rosaces. Je suis donc seul responsable de ce qui suit.
Bras
Un bras est une structure linéaire s’étendant entre deux points asymptotes (PA), qui sont des cœurs de rosaces, principale ou terminale. Chaque bras contient une infinité de microcardioïdes, parmi lesquelles une présente le poids minimum : cette microcardioïde est la majeure du bras, et par définition extensive le poids du bras est celui de sa microcardioïde majeure.
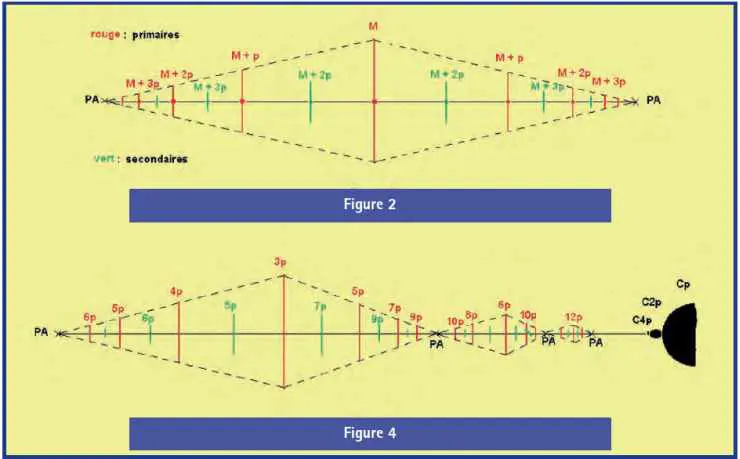
La répartition des microcardioïdes dans les bras libres est de type binaire : une cardioïde M au centre approximatif, encadrée de deux cardioïdes de poids M + p, quatre M + 2p, huit M + 3p, etc. (figure 2).
La taille et l’espacement des microcardioïdes du bras diminuent très rapidement quand le poids augmente, pour se stabiliser sur une loi logarithmique :
Log taille(A) – Log taille(B) = k (poids(B) – poids(A))
mais le coefficient k n’a de valeur que locale, déterminée au mieux pour un demi-bras, amont ou aval.
 Avec une telle loi, les cardioïdes M + kp, avec k = 0 à 5 pour fixer les idées, occupent la plus grande part de la longueur du bras, et les autres, vers les extrémités, ne deviennent visibles qu’en grossissant l’entourage des points asymptotes.
Avec une telle loi, les cardioïdes M + kp, avec k = 0 à 5 pour fixer les idées, occupent la plus grande part de la longueur du bras, et les autres, vers les extrémités, ne deviennent visibles qu’en grossissant l’entourage des points asymptotes.
Les poids des bras libres d’une rosace Rp sont tous différents : ces bras se partagent des poids p + 1 à 2p – 1. L’attribution des poids aux bras ne se fait pas dans l’ordre des numéros, mais selon une hiérarchie propre à la famille du cercle :
– le pas hiérarchique est 1 dans les séries primaires, c’est-à-dire que la hiérarchie s’y confond avec le numérotage,
– le pas hiérarchique est 2 dans les secondaires majeurs,
– pour tous autres cercles de poids m = ap + b, le pas hiérarchique est le coefficient a.
Les bras sont construits comme un enchaînement de maillons, qui dans les bras libres apparaissent au premier abord comme des triplets : deux rosaces majeures Rp encadrant une croix. La croix est définie comme une cardioïde portant deux rosaces latérales Rp, attachées par un bras latéral à ses cercles C3 (figure 3).
Les pédoncules ont une structure tout à fait particulière (figure 4) :
a) ils sont strictement rectilignes, du cercle au PA cœur de la rosace principale ;
b) ils ne sont pas symétriques longitudinalement. Leur majeure est 3p, et le pas aval est p comme dans les bras libres, mais le pas amont est 2p. Cette première section est le pédoncule propre du cercle Cp. À l’amont du pédoncule 3p, on trouve dans son prolongement un bras 6p, puis un bras 12p, etc. : ce sont les pédoncules des dessins associés aux cercles C2p, C4p, … qui existent nécessairement entre Cp et sa rosace, même si on ne les distingue pas à l’échelle de la rosace principale Rp ;
c) les maillons qui les constituent ne sont pas des triplets, mais seulement des croix. Les cardioïdes des pédoncules s’attachent l’une à l’autre directement, sans interposition de rosace.
Capsules
L’arborescence binaire des triplets d’un bras libre de rosace Rp se poursuit indéfiniment au pas p, mais elle n’en constitue pas l’unique structure. Si on examine le détail d’un bras, on observe que les rosaces majeures Rp du triplet séparent nettement celui-ci du reste du bras :
– à l’extérieur de la Rp du triplet M commence un bras de liaison qui s’étend jusqu’à la Rp du primaire voisin M + p. Ce bras est partagé par les deux rosaces et son poids est M + 2p. Sa description se déduit récursivement de celle du bras M et participe à sa structure (voir figure 2) ;
– entre les cœurs des deux Rp majeures se trouve la structure locale du triplet, que nous appellerons sa capsule et que nous allons détailler.
Toute cardioïde est encapsulée. Et, puisque la capsule contient de nombreuses microcardioïdes subordonnées, soulignons que cette affirmation est récursive. Moyennant quoi, nous pouvons réviser le schéma de principe du bras, tel que décrit plus haut, par quelques chiffres frappants, tirés d’un bras particulier typique :
– la capsule du triplet majeur occupe environ 45 % de la longueur du bras,
– l’ensemble des capsules de la série primaire de triplets représente 80 % de la longueur du bras,
– la longueur du plus grand bras secondaire, entre la capsule de M et celle de M + p, ne représente que 8 % de la longueur du bras, et non 25 % comme le laissait supposer le schéma de principe, avec ses microcardioïdes ponctuelles.
De tels chiffres montrent que la capsule ne peut être ignorée. Qu’il n’en soit parlé nulle part souligne que, depuis des années, les chercheurs ont abandonné les images pour revenir à l’abstraction.
La figure 5 montre l’organisation de principe de toute capsule, formée de couches imbriquées. Soit M le poids de la cardioïde centrale, triplet quelconque d’un bras de Rp.
Couche 1 : sur le même exemple, cette couche représente 64 % en longueur et 85 % en surface de l’encombrement total de la capsule.
Le long de l’axe d’approximative symétrie de la microcardioïde, la couche 1 se compose de deux structures de connexion amont et aval, composées chacune d’une des rosaces majeures (moins le bras de liaison), et d’un bras de connexion qui est d’ailleurs un des bras de la Rp. Ce bras de connexion s’étend du cœur de la Rp de couche 1 au cœur de la Rp de couche 2.
Si la cardioïde M appartient à un bras de numéro Bi, et si nous nommons B1 le bras de liaison du triplet vers l’extérieur, la rosace majeure Rp comporte p – 2 bras libres et son bras de connexion est le bras Bi. Les poids des bras libres de la Rp sont déterminés par les règles hiérarchiques normales en partant de B1 = M + p ; le bras Bi de connexion obéit à des règles particulières, décrites plus loin.
Sur la direction perpendiculaire, matérialisée par les cercles C3 de la microcardioïde M, la couche 1 comprend deux structures de remplissage droite et gauche (en regardant vers l’aval). Chacune de ces structures comprend une rosace latérale avec p – 1 bras libres, et un bras de remplissage qui descend vers la cardioïde, terminé par un point asymptote.
L’attribution de poids à ces divers bras obéit à des règles décrites plus loin. On retiendra pour le moment que les rosaces latérales sont approximativement symétriques par rapport à l’origine de la cardioïde.
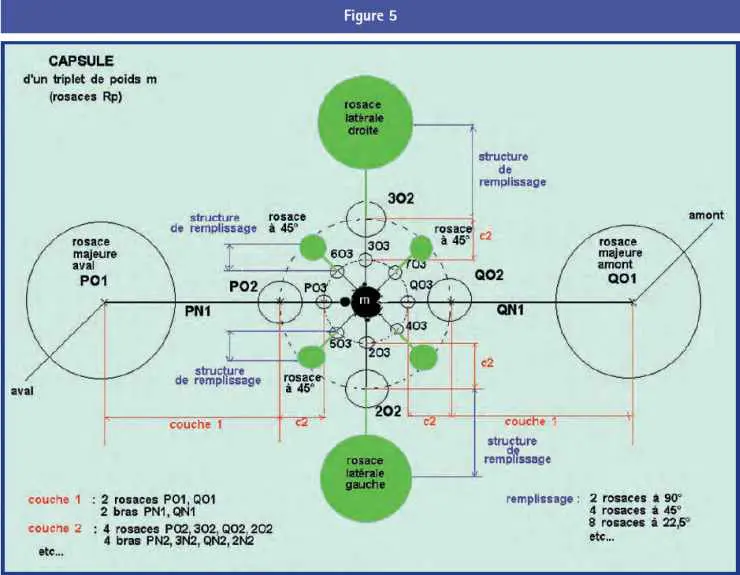
Couche 2 : très grossièrement, et simplement comme ordre de grandeur, la couche 2 occupe à l’intérieur de la couche 1 la même place relative que celle-ci dans l’ensemble de la capsule.
Elle comprend quatre structures de connexion identiques, déductibles l’une de l’autre par rotation d’environ 90° autour de la cardioïde centrale, dont les rosaces forment point asymptote intérieur pour les bras de connexion et les bras de remplissage de la couche 1.
Ces structures de connexion se déduisent par similitude des structures de couche 1, avec tous les poids augmentés de M. La relation logarithmique entre poids et taille explique la considérable diminution des dimensions, d’une couche à la suivante.

À 45° de ces quatre structures apparaissent quatre structures de remplissage, elles aussi déduites par similitude des structures de couche 1 avec poids augmenté de M. Ces structures émanent de points bien définis de la cardioïde, nommément les cercles C2.3 et C4.
Exemple figure 5A
Couche 3 : huit structures de connexion de poids encore accru de M s’attachent à l’intérieur des huit structures de couche 2. Huit structures de remplissage s’intercalent entre elles, émanant de huit points bien définis de la microcardioïde centrale.
Ce dispositif se poursuit indéfiniment, par doublement du nombre des structures à chaque couche. Des mesures ont été effectuées jusqu’à la couche 100 sur la cardioïde C2. B2MK de poids 3, la plus grosse de l’ensemble. Les points d’attache des structures de remplissage ont été déterminés jusqu’à la couche 13 (4 096 points d’attache propres).
Dès la couche 6, la forme des enveloppes cesse d’être quasi circulaire pour se modeler sur la forme de la microcardioïde, l’épaisseur des couches continuant à diminuer dans un rapport de l’ordre de 2,6 avec leur rang. De la sorte, il existe une limite à distance finie, que nous appelons la couche d’isolement : c’est l’ensemble de tous les points asymptotes terminant tous les bras de toutes les rosaces de la microcardioïde, homéomorphisme de la limite qui sépare l’Ensemble de la zone de divergence.
Structures de connexion – Évocation amont
L’exposé précédent montre qu’il suffit de connaître la structure de connexion de couche 1 pour connaître l’ensemble de la capsule. Le poids des cardioïdes du bras de connexion de couche 1 est déterminé par évocation amont, laquelle fonctionne comme suit :
Soit m le poids d’une cardioïde quelconque, primaire au sein d’un bras de poids M, délimité par deux points asymptotes dont l’un est forcément à l’amont de m ;
1) examiner toutes les cardioïdes entre m et ce PA amont, soit a le plus petit poids rencontré. a est le paramètre d’évocation amont de la cardioïde m, et le triplet majeur du bras de connexion de couche 1 aura un poids m + a ;
2) examiner toutes les cardioïdes entre m et a, soit q1 le plus petit poids rencontré, forcément > a. S’il existe plusieurs cardioïdes de poids q1 entre m et a, on choisira la plus proche de m. Le deuxième triplet du bras de connexion de couche 1 aura un poids m + q1 ;
3) recommencer cette recherche entre m et q1 pour trouver q2 > q1 et obtenir un triplet de couche 1 de poids m + q2. Cet exercice se poursuit tant que qi < a + m, car nous savons d’après les définitions que la couche 2 commence par un poids (m + a) + m ;
4) ainsi le bras de connexion contient un maximum de m cardioïdes primaires, de poids compris entre m + a et 2m + a – 1. L’ensemble de ces poids constitue le code caractéristique de la cardioïde m ; il est unique au sein du bras M ;
5) entre deux primaires m + qi et m + qi + 1 ainsi définis, il existe des secondaires de poids supérieur. Ce sont des évocations des secondaires situés entre les cardioïdes amont qi et qi + 1.
Si le bras M est un bras de rosace principale, toutes les cardioïdes susceptibles d’être utilement évoquées ont des poids de la forme M + k p. Les poids de couche 1 de m sont alors de forme m + a + i p.
Mais le principe d’évocation amont a une portée plus générale. Il s’applique à toute cardioïde m interne au bras M, par exemple une cardioïde d’un bras quelconque d’une rosace terminale, ou une cardioïde de capsule. L’exploration décrite plus haut franchira alors nécessairement un ou plusieurs PA, cœur de rosace terminale ou cœur de rosace de capsule. À chaque traversée de ce genre, la séquence au pas p est interrompue pour reprendre le calcul de la couche 1 avec une nouvelle origine.
Il est alors nécessaire d’introduire dans le bras de connexion une rosace intermédiaire, évocation de la rosace traversée dont elle recopiera la disposition des bras. Le code doit incorporer ces rosaces, et la capsule est dite feuilletée.
Le feuilletage est extrêmement répandu, et la structure des capsules se complique donc au fur et à mesure que l’on explore plus de détails. Cette propriété des structures mathématiques abstraites les distingue définitivement des structures du monde physique, qui finissent toujours – croit-on – par atteindre un niveau ultime, atomique au sens étymologique.
Structures de remplissage – Évocation aval
L’exposé sur les capsules a montré que la connaissance de la structure de remplissage de couche 1 suffit pour connaître toutes les structures de ce type dans la capsule.
Le poids des cardioïdes du bras de remplissage est déterminé par évocation aval. On ne reprendra pas le détail du procédé, qui examine cette fois l’intervalle séparant la cardioïde m du PA aval du bras M, et permet de définir un paramètre b d’évocation aval, qui n’a aucune raison d’être égal au paramètre a d’évocation amont, sauf si m = M.
L’évocation ne s’arrête pas au PA, cependant. Au-delà de ce PA, il existe dans le bras M une récursion illimitée de rosaces terminales, et la rosace latérale de m n’est autre que l’évocation de cette terminale. En résumé donc, l’ensemble de la structure de remplissage est l’évocation, par translation m de tous les poids, de tout ce qui existe dans le bras M à l’aval de la cardioïde m.
Conséquences
Les mécanismes exposés sont relativement simples, et je les ai appliqués pour une étude méthodique de deux bras parmi l’infinité des possibles :
– le grand bras C2. B2 de poids 3, qui est le bras libre unique de la rosace R2, émanation du cercle C2. L’étude visait à identifier individuellement toutes les cardioïdes de poids inférieur ou égal à 20, et à déterminer leur code caractéristique.
Elle s’est étendue aux terminales successives de poids 4 à 11, qui ont confirmé de façon particulièrement convaincante les prévisions sur le feuilletage des capsules.
– le bras CS5. B2 de poids 8, arbitrairement choisi parmi les quatre bras libres de la rosace du cercle secondaire CS5, en vue de contrôler l’universalité des principes découverts dans l’étude précédente. La limite de poids choisie est ici 50, car le pas est 5.
Cette étude a montré qu’il est parfaitement possible d’écrire un programme d’ordinateur pour déterminer automatiquement la structure d’un bras quelconque, de pas p et de poids M fixés, jusqu’à une profondeur (poids maximum) choisie a priori. Cependant, ce résultat numérique ne donne aucune idée de l’aspect réel des images d’un tel bras.
Je répète que toute cardioïde est encapsulée. On est donc conduit à réfléchir sur la structure de capsules imbriquées.
Ce travail a été poussé jusqu’à ses ultimes conséquences, et a permis d’expliquer de nombreuses observations a priori bizarres. Je me borne à les énoncer ; le lecteur curieux pourra soit consulter les références in fine, soit m’aider à organiser une présentation de diapositives ou un cours sur ordinateur :
– amas de cardioïdes résultant d’évocations récursives (figures 6 et 7),
– convergence vers une structure commune de triplets de couches profondes dans des capsules correspondant à des valeurs différentes de p (figure 8),
– structure remarquable des carrefours dans les arborescences de tels triplets (figure 9),
– apparition de structures spécifiques, récursives et susceptibles de se combiner, à partir du deuxième niveau d’encapsulation. Je les ai baptisées boules, d’après leur forme (figure 10).
Cas des B1 pédoncules
Puisqu’il n’y a pas de rosace dans les maillons des bras B1 pédoncules, que devient dans ces bras le concept de capsule ?
a) L’évocation amont s’applique intégralement, et porte exclusivement sur des cardioïdes de bras B1, par construction. Mais puisque aucune rosace ne risque de s’interposer et de couper les séquences de poids, il est permis qu’une cardioïde particulière joue deux rôles, pourvu que son poids l’autorise :
– membre de l’arborescence M + k p des cardioïdes structurelles du bras,
– élément d’une couche de capsule.
Cette interpénétration est assez fréquente et complique l’analyse. Elle réduit notablement le nombre de cardioïdes de poids donné dans un bras B1, par rapport à un Bi de même poids.
b) L’évocation aval s’applique également : toutes les rosaces latérales des B1 pédoncules sont donc des évocations de la seule rosace visible à leur aval, c’est-à-dire la rosace principale.
c) Les bras B1 des cercles C2p, C4p, etc., en amont du B1 de Cp, évoquent cette même rosace : quel que soit le cercle qui leur correspond, les croix de ces B1 ont donc des rosaces latérales à p bras.
Cercles de niveau supérieur à 1
Considérons un cercle Cp tangent à la cardioïde de base, portant un cercle Cq qui porte lui-même un cercle Cr. On a déjà indiqué que le poids de ce dernier cercle est p q r. Que sont les dessins émanant du cercle Cp.q.r ?
Ce sont des rosaces arborescentes qui expriment, dans l’ordre inverse, les rangs de tous les cercles traversés :< ul>
- une rosace de base Rr à r bras dont un pédoncule de poids p q r , et r – 1 bras libres de poids k1n1 = p q (r + n1) ;
- sur chaque bras libre de Rr, une rosace Rq à q – 1 bras libres, à laquelle le bras de la Rr sert de B1.
Le poids des bras de la Rq issue du bras majeur de Rr est
k21n2 = k11 – 2pq + p n2.
Le poids des autres bras est
k2n1 n2 = k1n1 – pq + p n2.
Le bras majeur est donc beaucoup plus gros que les autres. - sur chaque bras libre de chaque Rq, une rosace Rp à p – 1 bras libres, à laquelle le bras de la Rq sert de B1. Le poids des bras de la Rp issue du bras majeur de la Rq est
k3n1.1n3 = k2n1.1 – 2p + n3.
Le poids des autres bras est
k3n1n2n3 = k2n1n2 – p + n3.
La rosace du bras majeur est donc plus grosse que les autres ; - et bien entendu tous les bras libres de ces Rp s’achèvent sur une terminale récursive telle que
k4n1n2n3 = k3n1n2n3 + n4.
 L’effet de ces formules est que le dessin apparaît à première vue comme une seule rosace Rp de poids k311n3, bien plus grosse que toutes les autres rosaces que l’observation attentive peut ensuite reconnaître.
L’effet de ces formules est que le dessin apparaît à première vue comme une seule rosace Rp de poids k311n3, bien plus grosse que toutes les autres rosaces que l’observation attentive peut ensuite reconnaître.
Bien plus compliquée que la structure des dessins de premier niveau, celle des dessins de niveaux supérieurs a été complètement éclaircie, quel que soit le niveau, et résulte simplement d’une application sans concession des lois précédemment découvertes. Néanmoins, il n’est pas réaliste de les présenter ici.
Voir un exemple de rosace de 3e niveau en figure 11.
Noyaux
La description précédente, à base de bras de rosaces, ne concerne que les structures majeures obtenues en plaçant bout à bout des cardioïdes. On a bien fait état des rosaces latérales et de leur interprétation, mais elles n’occupent que peu de place dans les dessins d’ensemble.
L’examen détaillé de ces structures latérales permet cependant des découvertes remarquables, en ce que leurs images combinent les dispositifs caractéristiques du bras avec les conséquences de la traversée d’un cercle Cn subordonné à la microcardioïde. L’effet est de remplacer chaque microcardioïde par une structure complexe baptisée noyau.
Les lois de formation des noyaux, qui combinent plusieurs niveaux d’évocation, ont été complètement déterminées. Il s’agit d’images exceptionnellement chargées, et souvent fort belles, mais il est impossible de s’étendre sur ce vaste sujet dans l’espace disponible ici.
Le lecteur doit cependant être conscient que ces noyaux sont partout : en particulier, tous les triplets de bras latéraux sont des noyaux de C3.
Voir figure 12 pour un exemple.
Articles similaires :
 Quelques souvenirs d’un petit colon français au Tonkin (Viêtnam)…
La production informatique, élément majeur du fonctionnement des entreprises
Quelques souvenirs d’un petit colon français au Tonkin (Viêtnam)…
La production informatique, élément majeur du fonctionnement des entreprises
 Thomas Jefferson et l’École polytechnique : à la recherche des chaînons manquants
Thomas Jefferson et l’École polytechnique : à la recherche des chaînons manquants
 Épidémiologie environnementale
Alfred Grosser : réflexion sur le lien franco-allemand
Épidémiologie environnementale
Alfred Grosser : réflexion sur le lien franco-allemand