Biomédecine et physique : Mouvements collectifs de micronageurs confinés

L’essor de nouvelles techniques de microscopie et de micromanipulation a permis dans les dernières décennies des avancées spectaculaires dans l’étude des nageurs microscopiques, en particulier celle des bactéries. De nombreuses questions persistent cependant quand l’environnement de ces nageurs est complexe, par exemple en présence de parois ou d’autres nageurs. Le présent travail allie expériences et théorie pour analyser le comportement d’une suspension confinée de bactéries magnétotactiques, avec des applications en biologie et en ingénierie médicale.
Dans Le Voyage fantastique, film de science-fiction de 1966, le sous-marin Proteus et son équipage sont miniaturisés et injectés dans le corps d’un scientifique : ils ont une heure pour détruire un caillot dans son cerveau et le sauver.
Micronageurs : de la fiction à la médecine ?
Les effets spéciaux qui illustrent le monde microscopique de notre organisme et les dangers qui y rôdent ont valu à l’équipe du film deux Oscars. Mais qu’en diraient un médecin, un biologiste ou un physicien ? Quelques heures dans les salles obscures ont déjà pu suffire à vous convaincre qu’Hollywood ne va pas de pair avec la rigueur scientifique. Une telle miniaturisation, en particulier, est irréalisable : en physique, la taille caractéristique d’un système compte. On ne peut pas réduire ou grossir arbitrairement un objet tout en conservant ses propriétés et son fonctionnement. Ainsi, et nous y reviendrons, les lois qui régissent les fluides à l’échelle macroscopique et à l’échelle microscopique ne sont pas les mêmes. Et pourtant : l’idée d’injecter dans le corps d’un patient un nageur microscopique contrôlé de l’extérieur pour des opérations moins invasives que par des moyens classiques n’est pas réservée aux producteurs de cinéma. Elle est l’objet d’une recherche biomédicale pluridisciplinaire dont les applications envisagées sont nombreuses. Comme dans Le Voyage fantastique, on imagine déloger les caillots sanguins, mais aussi réaliser des chirurgies artérielles plus complexes.
Si le nageur peut transporter des médicaments jusqu’à une zone déterminée, des traitements locaux pourraient être élaborés, notamment pour s’attaquer de façon ciblée aux tumeurs cancéreuses. De tels robots sont déjà développés dans de nombreux laboratoires et leurs concepteurs cherchent à les rendre plus petits, plus précis, plus contrôlables. Pourtant, leur réalisation soulève de nombreuses questions : quelle sera la réponse du système immunitaire à leur présence ? Comment les fabriquer de manière reproductible et peu coûteuse ? Après leur utilisation, comment seront-ils dégradés ou évacués ? Leur motilité (i.e. leur capacité à se déplacer continûment dans un liquide en déformant leur corps) pose en particulier des problèmes fondamentaux, abordés dans la présente thèse : comment parviendraient-ils à se déplacer, s’orienter, éviter les obstacles et interagir efficacement avec d’autres nageurs ?
De la difficulté de nager à l’échelle microscopique
Pour mieux appréhender les enjeux de la nage à l’échelle microscopique, nous devons revenir à la différence, évoquée plus haut, entre un fluide à l’échelle macroscopique, celle des océans, des lacs, des piscines, mais aussi d’un verre d’eau, et un fluide à l’échelle microscopique, celle d’une goutte d’huile, de nos capillaires sanguins, des cellules. La première nous est bien plus familière ; et pourtant c’est dans la deuxième qu’évoluent la plupart des êtres vivants de la pièce où vous vous trouvez.
En général, le comportement d’un fluide est régi par l’inertie et la viscosité. La première correspond au fameux terme « masse fois accélération » des équations de Newton, qui rend la chute d’une pomme sur la tête dudit Newton plus douloureuse si le pommier est haut. La viscosité est une grandeur plus élusive, qui mesure la résistance d’un liquide à l’écoulement : l’acétone ou l’eau ont une viscosité faible, alors que celle de la glycérine ou de la résine est élevée. Le nombre de Reynolds permet de comparer l’importance relative de l’inertie et de la viscosité.
Pour l’évaluer, considérons un nageur de taille L, qui se déplace à une vitesse U, dans un fluide caractérisé par sa densité ρ et sa viscosité µ. Le nombre de Reynolds est le ratio adimensionnel des forces inertielles et visqueuses : Re = ρUL/µ. Dans le monde microscopique, U et L étant très petites, ce nombre de Reynolds est très inférieur à un et l’inertie disparaît : l’environnement est entièrement régi par les forces visqueuses. On notera qu’un très petit nombre de Reynolds peut aussi exister à de très grandes tailles : il suffit pour cela que la viscosité du liquide soit très élevée. C’est le cas dans le manteau terrestre, dont la viscosité est 1024 plus élevée que celle de l’eau ! Les forces gravitationnelles de la Terre ont beau être immenses, l’écoulement du manteau reste très lent et dominé par la viscosité.
Le nombre de Reynolds pour un nageur dans une piscine est de l’ordre de 104 ; pour les micronageurs biologiques que nous étudions, comme les bactéries et les spermatozoïdes, il est de 10-4. Alors, à quoi ressemble la vie à très bas nombre de Reynolds ? Edward Mills Purcell, dans une conférence de 1977, proposait l’analogie suivante : vous êtes dans une piscine de mélasse, et de plus vous ne pouvez bouger votre corps qu’à 1 cm/min, si bien que vos mouvements de bras ne dépassent pas la vitesse des aiguilles d’une horloge. Si dans ces conditions vous parvenez à vous déplacer de plusieurs mètres en quelques jours, vous êtes bien un nageur à bas nombre de Reynolds.
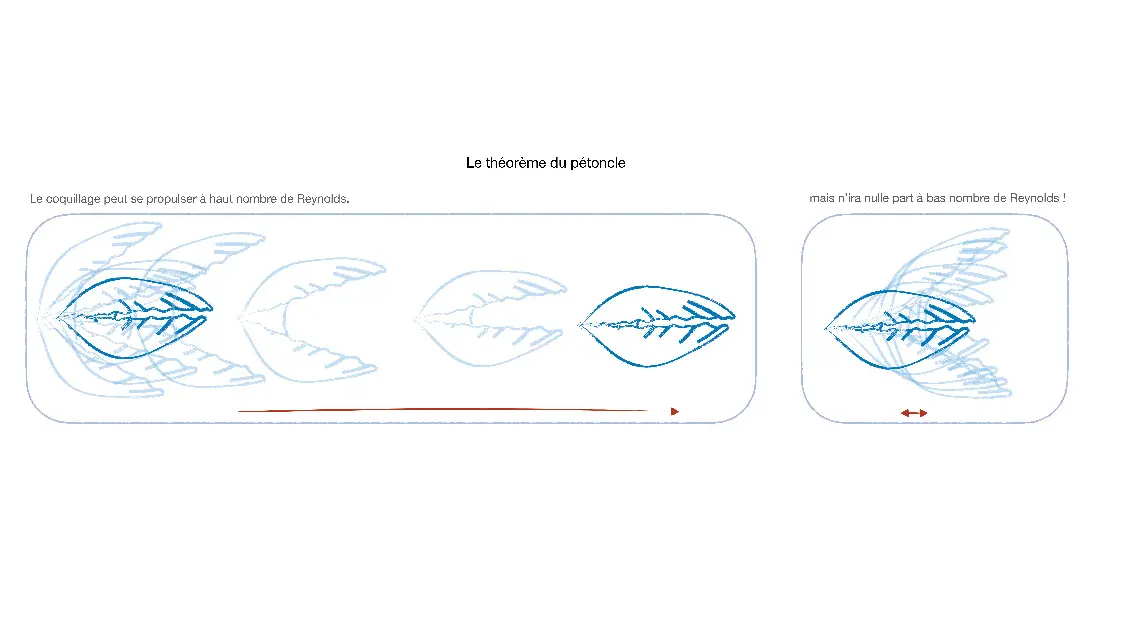
La performance n’est pas anodine, puisque l’inertie représente la capacité à avancer encore quand nous arrêtons de bouger, par exemple après la poussée contre le mur d’une piscine. Elle est inexistante pour une bactérie : si celle-ci arrête de nager, elle s’immobilise après 0,1 angström – la taille d’un atome. Son déplacement est entièrement déterminé par son action présente, et par rien de ce qui est arrivé dans le passé : la nage est instantanée. Se mouvoir est d’autant plus difficile que l’absence d’inertie a une conséquence additionnelle : un mouvement réversible (i.e. identique si le temps avance ou remonte) ne peut conduire à aucun déplacement. Aucune modulation de rythme ne peut y remédier : cette propriété est surnommée « théorème du pétoncle » et est illustrée ci-dessous. Ni le battement des nageoires, ni celui des ailes, ni le mouvement d’une coquille ou l’expulsion de l’eau du corps d’une méduse ne permet de quitter le point de départ.
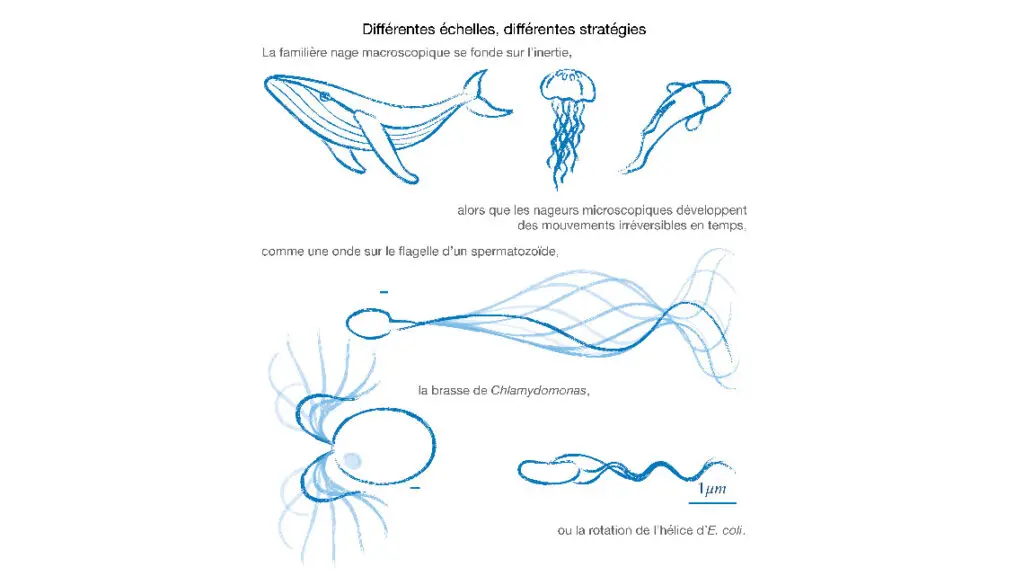
Stratégies de nage microscopique
Une stratégie de nage à l’échelle macroscopique sera inapplicable à l’échelle micrométrique. Mais alors, comment se déplacer ? L’image ci-après donne quelques exemples de nage microscopique, dont le point commun est de briser la symétrie entre le mouvement exécuté à l’endroit, vu en marche avant, et à l’envers, en marche arrière. Certains planctons utilisent des flagelles qu’ils déforment en mouvements non réversibles, comme la brasse de l’algue verte Chlamydomonas reinhardtii. Les spermatozoïdes, eux, exploitent l’asymétrie d’une onde qui se propage de leur queue vers leur tête. Les bactéries, enfin, possèdent un ou plusieurs appendages rigides, en forme d’hélice, dont la rotation dans le sens contraire des aiguilles d’une montre permet un mouvement net. L’efficacité de la nage bactérienne est remarquable : E. coli, quand elle nage, se déplace à plus de 20 µm/s, soit environ quatre fois sa taille à chaque seconde, plus vite relativement qu’un nageur olympique ou même un dauphin.
L’étude des nageurs microscopiques, en particulier celle des bactéries, a pris son essor dans les années 2000 grâce à l’avènement de techniques de microscopie qui ont permis d’observer pour la première fois les flagelles de bactéries. Si leur nage en milieu homogène est depuis lors bien caractérisée, de nombreuses questions persistent quand l’environnement est plus complexe, par exemple en présence d’autres nageurs ou de parois. Or la plupart des micro-organismes évoluent dans de tels milieux, comme l’humus terrestre ou le corps humain pour les applications médicales. L’objet de ma thèse est justement de proposer des modèles valides pour les micronageurs confinés, en particulier pour les suspensions, qui désignent un grand nombre de nageurs dispersés dans un même liquide. La suite de cet article présente une partie de ce travail, qui s’est concentré sur des suspensions de bactéries dites magnétotactiques.
Se diriger grâce aux champs magnétiques
Ces bactéries ont une propriété singulière : elles s’orientent en suivant les champs magnétiques de leur environnement. En effet, elles possèdent dans leur corps des nano-aimants, appelés magnétosomes. Un champ extérieur crée un moment qui aligne ces aimants et donc le corps de la bactérie, avec lui. La force exercée étant négligeable, la vitesse de la bactérie est contrôlée par sa nage, mais elle est orientée par le champ extérieur. Une possible explication à ce comportement est l’utilité pour une bactérie de se guider en utilisant le champ magnétique terrestre, puisque la gravité est négligeable et que les bactéries ne peuvent distinguer le haut du bas ou de toute autre direction dans leur environnement.
Le magnétotactisme est prometteur pour de nombreuses applications puisqu’il permet de guider un nageur avec un champ inoffensif pour un être humain, soit en utilisant directement les bactéries, soit en conférant la même propriété à un nageur artificiel. Il modifie cependant de manière significative le comportement des nageurs, tant pour un nageur isolé que pour une suspension, ce qui requiert un travail spécifique pour modéliser et prédire leur comportement. Par exemple, puisqu’ils sont guidés par le champ magnétique plutôt que par les propriétés locales de leur environnement, ils ont tendance à s’accumuler contre des obstacles et des parois.
Comment des bactéries magnétotactiques interagissent-elles avec leur environnement ?
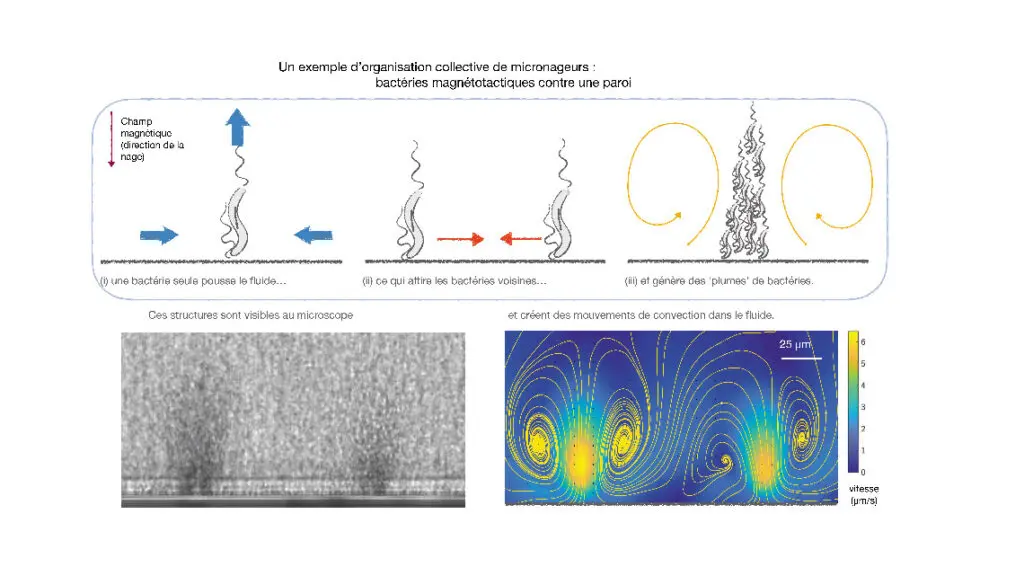
Notre travail allie des expériences et un modèle pour analyser le comportement d’une suspension de bactéries magnétotactiques. De nombreuses bactéries sont introduites dans un capillaire en verre à section carrée, puis nous générons un champ magnétique perpendiculaire à l’axe du tube. Ce dispositif nous permet d’observer l’accumulation de bactéries magnétotactiques contre l’une des parois et de mesurer la circulation du fluide dans le capillaire. Plutôt qu’en une couche uniforme de bactéries qui nageraient perpendiculairement au mur, nous observons rapidement la formation de « plumes » perpendiculaires à la paroi. Ces plumes créent des recirculations semblables à de larges cellules de convection, qui grandissent jusqu’à atteindre la taille du capillaire.
“Le magnétotactisme est prometteur pour de nombreuses applications.”
Ce phénomène d’accumulation locale de nageurs peut être décrit par un modèle hydrodynamique simple. Lorsqu’elle nage contre une paroi, une bactérie ne peut pas bouger et se contente alors d’expulser du fluide derrière elle. Tout se conserve, en particulier la masse du fluide environnant : cette expulsion vers l’arrière est compensée par l’arrivée de liquide sur les côtés, comme sur le schéma ci-contre. Lorsque plusieurs bactéries poussent ainsi contre le mur, elles attirent donc le fluide autour d’elle et, avec lui, leurs voisines. De petits groupes se forment et se regroupent à leur tour. Lorsqu’ils atteignent une taille critique, le fluide qu’ils expulsent vers l’arrière entraîne des bactéries, d’où la formation de plumes allongées perpendiculaires au mur.
La circulation de fluide déplacé par la nage des bactéries crée en retour les structures que nous observons dans le fluide. Cette attraction entre bactéries voisines et l’organisation de la suspension qui en découle ne peuvent avoir lieu qu’en présence du champ magnétique. Si le champ est éteint, ou que les bactéries ne s’alignent pas, la solution reste uniforme et désordonnée. Notre travail a ainsi permis de montrer que l’existence d’un mécanisme d’orientation externe aux nageurs, magnétique ou non, transforme radicalement l’interaction des bactéries avec les parois de leur environnement.
À l’échelle microscopique, les petits détails comptent
D’autres expériences observaient les bactéries dans une goutte sphérique plutôt que contre une paroi. Bien que les deux systèmes soient très proches, leurs comportements sont très différents : les bactéries créent un vortex d’axe vertical et la goutte se met à tourner dans le sens des aiguilles d’une montre. C’est cette dernière propriété, une rotation avec une direction définie, qui nous a le plus intrigués : pourquoi tourner dans ce sens et non parfois dans le sens horaire et parfois dans l’autre, comme le prédisait le modèle que nous avions développé ? Nous avons alors montré que la courbure des parois de la goutte permettait d’observer un autre phénomène, jusqu’alors négligé : la chiralité du flagelle de la bactérie.
La chiralité, du grec kheir – main, est la propriété d’un objet qui n’est pas superposable à sa réflexion dans le miroir : une main ou, ici, une hélice. Cette chiralité est transmise, de par la rotation de l’hélice, au fluide autour d’une bactérie ou d’un groupe de bactéries, et influe sur leur interaction avec une paroi courbe ou inclinée. La rotation de tous les flagelles dans le même sens, couplée à la géométrie de l’environnement et à la gravité, est finalement transmise à la goutte entière et la fait tourner dans le sens horaire. Cette propriété microscopique devenue dominante à l’échelle de la goutte doit donc être prise en compte dans nos modèles de nage dans des géométries complexes. À cette échelle, même les plus petits détails comptent !
En somme, notre travail a permis de mieux comprendre le rôle d’une orientation externe dans le comportement de groupes de micronageurs, avec des applications possibles en biologie et en ingénierie médicale. Plus largement, il s’agit d’un exemple des problèmes interdisciplinaires très riches qui se posent en biologie et peuvent être abordés avec les outils de la physique et de la mécanique classique. Réciproquement, le monde vivant génère une profusion de nouvelles questions en mécanique des fluides, avec un cadre très différent des questions classiques d’ingénierie, comme ici les fluides à très petites échelles. C’est sur ces questions que portent l’ensemble de ma thèse et ma recherche actuelle.
Ressources
- E.M. Purcell, Life at low Reynolds number, American Journal of Physics 45,3 (1977). https://doi.org/10.1119/1.10903
- B. Vicenti, G. Ramos, M.L. Cordero, C. Douarche, R. Soto et É. Clément, Magnetotactic bacteria in a droplet self-assemble into a rotary motor, Nature communications 10, article n° 5082 (2019). https://doi.org/10.1038/s41467-019–13031‑6
- A. Théry, L. Le Nagard, J.-C. Ono-dit-Biot, C. Fradin, K. Dalnoki-Veress, E. Lauga. Self-organisation and convection of confined magnetotactic bacteria. Scientific Reports 10, article n° 13578 (2020). https://doi.org/10.1038/s41598-020–70270‑0
Informations sur la thèse
Le travail présenté ici a été réalisé dans le cadre d’une thèse en mathématiques appliquées à l’Université de Cambridge intitulée Confinement-mediated accumulation and collective dynamics of microswimmers (Accumulation et organisation collective de micronageurs confinés). La thèse a été soutenue en mai 2022, le jury de thèse était composé de Raymond E. Goldstein (Université de Cambridge) et Cécile Cottin-Bizonne (Institut Lumière Matière, Lyon).
https://doi.org/10.17863/CAM.85810
Laboratoire d’accueil
Cette thèse a été réalisée dans le groupe d’Éric Lauga au DAMTP (Département de Mathématiques Appliquées et Physique Théorique) de l’Université de Cambridge au Royaume-Uni. Elle a été financée par une ERC de l’Union Européenne attribuée à Éric Lauga dans le cadre du programme Horizon 2020. L’activité de recherche du groupe est centrée sur le développement de modèles quantitatifs pour les nouvelles questions en mécanique des fluides biologiques, grâce à une combinaison de modèles analytiques, d’analyses asymptotiques et de simulations numériques. Les problèmes étudiés incluent la locomotion dans des fluides visqueux, ainsi que la modélisation des fluides biologiques à l’échelle cellulaire, et de multiples problèmes d’hydrodynamique physique à petite échelle.
https://www.damtp.cam.ac.uk/user/lauga/index.html
Les expériences décrites ci-dessus ont été réalisées dans le laboratoire de Kari Dalnoki-Veress, au Département de Physique et Astronomie de l’Université McMaster, au Canada. Elles ont été réalisées par AT, Lucas Le Nagard, Jean-Christophe Ono-dit-Biot, sous la supervision de Cécile Fradin et Kari Dalnoki-Veress.
Articles similaires :
 Savant cherche refuge. Comment les grands noms de la science ont survécu à la Seconde Guerre mondiale
Savant cherche refuge. Comment les grands noms de la science ont survécu à la Seconde Guerre mondiale
 Hommage à Claudine Hermann : une pionnière pour les femmes en sciences
Hommage à Claudine Hermann : une pionnière pour les femmes en sciences
 Faire progresser le monde de la santé
Faire progresser le monde de la santé
 L’accès aux données de santé : un enjeu de souveraineté et d’attractivité pour la France et l’Europe
L’accès aux données de santé : un enjeu de souveraineté et d’attractivité pour la France et l’Europe
 CorWave, la medtech qui révolutionne les pompes cardiaques
CorWave, la medtech qui révolutionne les pompes cardiaques